
Характеристикой биологии, которая поразила меня за эти годы, является явление рецидивирующей логики проектирования, даже в системах, которые не связаны с происхождением. Это довольно удивительная характеристика в случае, если безумный процесс отвечает за происхождение жизни, но это именно то, что мы могли бы предсказать в гипотезе, где дух сыграл важную роль. В других областях опыта, когда мы встречаем логику повторяющегося дизайна, мы обычно связываем его с умными причинами. Например, существуют характеристики картин, которые характеризуют работу конкретного художника, и характеристики зданий, которые являются общими для зданий, разработанных тем же архитектором. Есть даже аспекты письма, которые отличаются от отдельного автора. Поэтому это пример прогнозирования, сделанный гипотезой о преднамеренной концепции — то, что мы ожидаем увидеть, предполагая участие интеллектуального духа. Есть много примеров этого типа явления в биологии. В целях иллюстрации я сосредоточусь здесь на классе рецидивирующей логики проектирования: две системы регуляции в бактериях.
Каковы две системы регулирования?
Бактериальные клетки используют системы с двумя компонентами для обнаружения и реагирования на изменения окружающей среды. Как следует из их названия, две системы, как правило, включают в себя два компонента: датчик киназы и регулятор отклика. Датчик киназа, в ответ на химический или физический стимул, подвергается аутофосфорилированию, с помощью которого фосфатная группа переносится из АТФ в остаток гистидина на киназе. Гистидиновая протеинкиназа имеет две области: площадь входа и поле выпуска. Первый расположен за пределами ячейки и идеально расположен для обнаружения входящих сигналов окружающей среды. Последний расположен на цитоплазматической стороне клеточной мембраны и расположена так, чтобы они могли взаимодействовать с регулятором ответа. Группа фосфатов переносится в регулятор ответа, который затем пилотирует клеточный ответ, такой как активация или дезактивация генов.
Две компонентные системы чрезвычайно распространены среди бактерий, и каждая использует одну и ту же основную логику проектирования. Далее я дам краткий обзор некоторых примеров этого типа.
Регуляция внешних мембранных белков в Escherichia coli
Двухконтранентная система регулирует экспрессию порина в ответ на осмолярность окружающей среды (мера концентрации растворенных частиц в растворе). Датчик киназа этой системы, расположенная во внутренней мембране, является Envz. Envz обнаруживает изменения осмолярности и подвергается аутофосфорилированию. Регулятор ответа, OMPR, получает фосфатную группу Envz и регулирует экспрессию генов.
Когда осмолярность высока, киназа активности envz активируется, что приводит к фосфорилированию OMPR. Когда осмолярность низкая, активируется фосфатазная активность ENVZ, снижая фосфорилированные уровни OMPR.
Во время фосфорилирования OMPR становится активным димером с улучшенной способностью связи ДНК, специфичной для Oppc И OMMPF генные промоутеры. Это пориновые гены, которые кодируют белки внешней мембраны (которые позволяют проходить метаболиты через внешнюю мембрану граммогативных бактерий). Диаметр пор OMPF больше, чем у OMPC. Это позволяет диффузию в десять раз быстрее, что выгодно в условиях низкой осмолярности, где питательные вещества редки. Если осмотическое давление низкое, синтез OMPF увеличивается. Если осмотическое давление высокое, экспрессия OMPC увеличивается. Кроме того, транскрипция МикрофРНК Antissens инициируется. Микроф блокирует перевод OMMPF Пополнительным соединением — синтез OMPF, таким образом, подавляется.
Хемотаксис регулирование
Бактерии способны перейти к источнику пищи, такому как глюкоза, с помощью процесса, называемого «хемотаксисом». Предварительным условием для работы этого процесса является способность бактериального жгутикового двигателя буквально изменять скорость, чтобы перейти от вращения в противоположном направлении игл от часов к вращению в направлении игл часов. Это изменение вращения вызвано в ответ на химические стимулы из внешней части клетки. Эти химические сигналы обнаруживаются с помощью схемы трансдукции сигнала с двумя компонентами, которые работают, индуцируя изменение вращения флагельера.
Читатели могут найти полезным обращаться к следующей диаграмме при чтении следующих описаний:
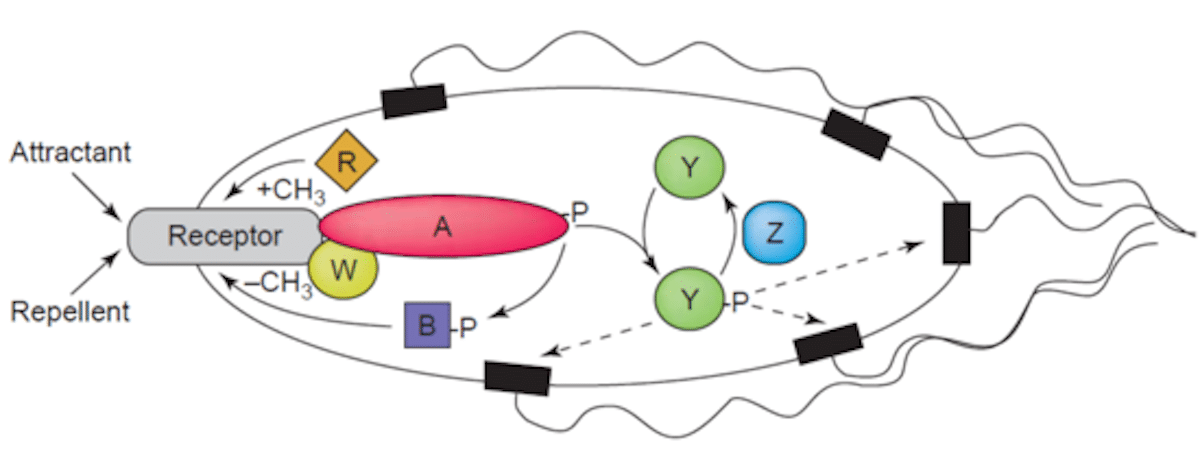
Как бактерии обнаруживают химический градиент? Ответ заключается в определенном классе трансмембранных рецепторов, называемых белками хемотаксиса, принимающими метил (далее MCP). Различные MCP могут обнаруживать различные типы молекул и способны связывать привлекательные или репелленты. Затем эти рецепторы общаются с — и активируются — так называемые «Che» белки ».
Белки, называемые Chea и Chew, связаны с приемником. Первая — гистидинкиназа этой системы. При активации приемника остаток гистидина, созданный KEA, подвергается самоуверенному. Есть два регулятора ответа под названием Cheb и Chey. Существует перенос группы фосфорила в их остаток аспартата Chea. Затем Чей взаимодействует с белком, переключающим флагелл, называемый Flim. Это вызывает переключение в жгутиковом направлении противоположного направления игл от часов к направлению игл часов.
Это вращение в направлении игл общения расстраивает весь луч жгутиков и вызывает его разрыв. Результатом является то, что бактерии «падают». Это означает, что бактерии способны перенаправить свою траекторию и переоценить и несколько раз корректировать свои ориентации в ответ на экологические стимулы, такие как пища или яды.
Что касается другого регулятора ответа, который я упомянул, Cheb, что он делает? Когда CHEB активируется гистидинкиназой Chea, он работает как ме истекает ме истекает. Это означает, что он активно устраняет метильные группы остатков глутамата на цитоплазматической поверхности приемника. Между тем, другой белок (называемый дорогой) активно добавляет метильные остатки в эти же остатки глутамата, то есть он работает как метилтрансфераза.
На этом этапе инженерная инженерия является гениальным ходом. Если стимул находится на высоком уровне, произойдет соответствующее снижение уровня фосфорилирования белка CHEA — и, следовательно, регуляторы ответа Chey и Cheb также. Помните, что роль Cheb состоит в том, чтобы устранить метильные группы из остатков глутамата, присутствующих на цитоплазматической поверхности приемника. Но в настоящее время фосфорилированный Cheb недоступен, и поэтому эта задача не выполнена. Степень метилирования MCP будет увеличена. Когда MCP полностью метилируются, клетка непрерывно плавает, потому что MCP больше не реагируют на стимулы.
Это подразумевает, что уровень фосфорилатов Chea и Cheb будет увеличиваться, даже когда привлекательный уровень останется высоким, и что ячейка начнет процесс всасывания. Но теперь фосфорилированный Cheb способен к Demethyler MCP, и рецепторы снова способны реагировать на привлекательные химические сигналы. В случае репеллентов ситуация похожа, за исключением того, что это наименее метилированные MCP, которые реагируют наименьшее, в то время как полностью метилированный MCP реагирует больше всего. Этот тип регуляции также означает, что бактерии имеют систему памяти для химических концентраций недавнего прошлого и сравнивают их с сигналами, которые она в настоящее время получает. Таким образом, он может обнаружить, приближается ли он или уходит от химического стимула.
Обнаружение кворума
Цель определения кворума по существу состоит в том, чтобы достать достаточное количество клеток данного вида, прежде чем запустить ответ, который требует, чтобы плотность популяции превышала определенный порог. Единственная бактериальная клетка, выделяющая токсин в эвкакариотическом организме, вряд ли повредит хозяину и отпускает ресурсы. Однако, если все бактериальные клетки большой популяции координируют экспрессию токсина, это, скорее всего, окажет желаемый эффект.
Каждый вид, который использует обнаружение кворума, которое включает в себя большинство грамотрицательных бактерий, а также определенные грамположительные бактерии синтезирует крошечную сигнальную молекулу (технически называемую «самостоятельно»), которая свободно распространяется через клеточную мембрану. Самостоятели специфичны для вида, что означает, что каждая клетка одного и того же вида производит одну и ту же молекулу. Это означает, что освобожденного человека не присутствует в высоких концентрациях внутри ячейки только тогда, когда многие клетки одного и того же вида находятся поблизости. Внутри клетки самооценка связывается с специфическим активирующим белком этой конкретной молекулы и, таким образом, сигнализирует о бактериях, чтобы начать транскрипцию специфических генов.
Взять, к примеру, случай биолюминесцентных бактерий, Аливибрио Фишери (Фото вверху этой статьи). Свет, который этот вид проблем бактерий является результатом действия энзийма люцифразы. Активирующий белок, называемый LUXR, отвечает за контроль операций LUX, которые, в свою очередь, ответственны за транскрибирование белков, необходимых для люминесценции. Эти операции индуцируются при концентрации конкретной самостоятельной занятости Аливибри Рыбы достиг достаточно высокой концентрации. Эта самозанятость сама синтезируется ферментом, который кодируется геном Luxi.
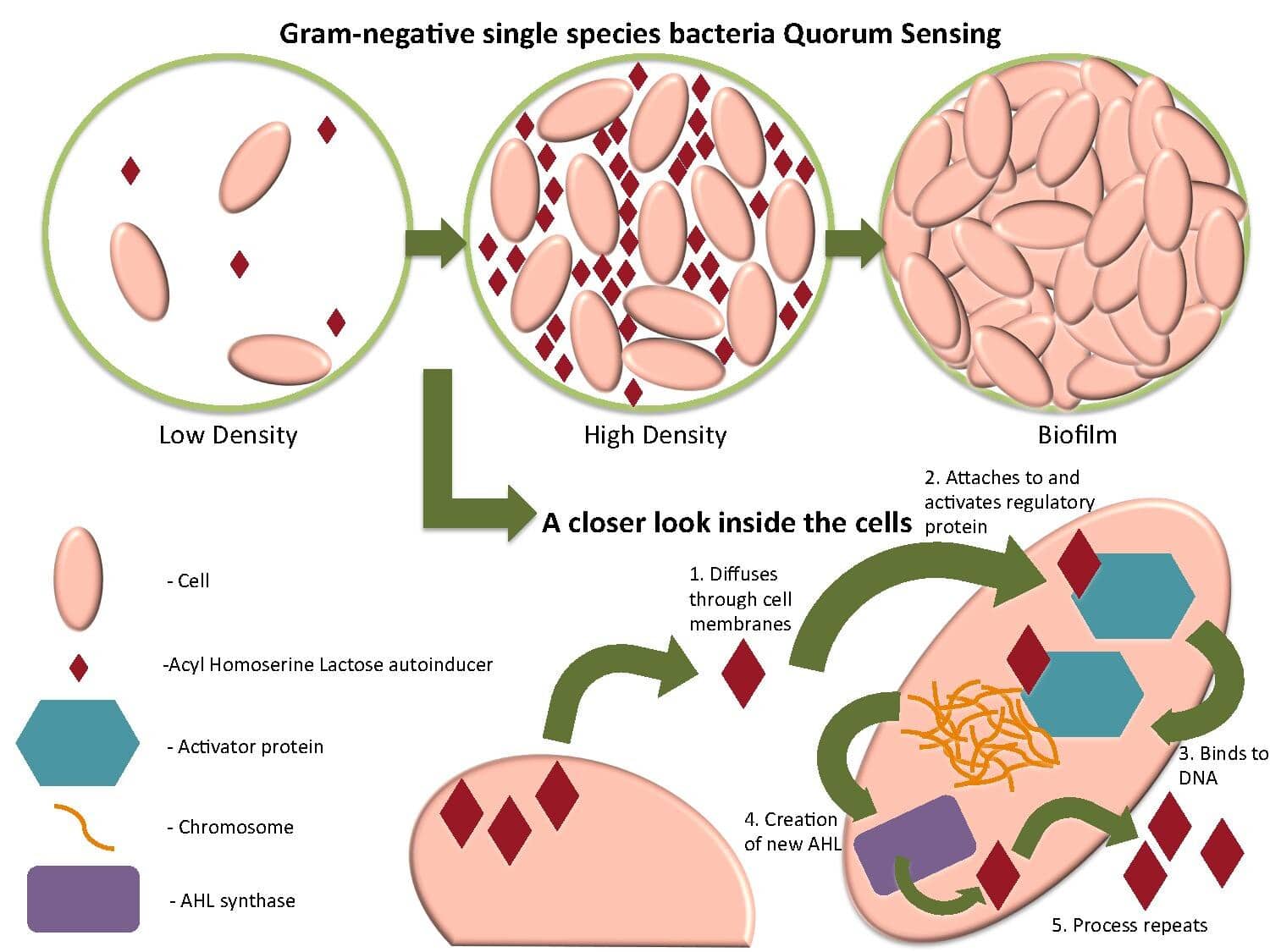
Ощущение кворума очень широко распространено, особенно у грамогативных бактерий. Pseudomonas aeruginosaНапример, использует такие процессы «отбора проб населения», чтобы вызвать экспрессию большого количества неродственных генов, когда плотность популяции достигает определенного порога. Эти гены затем позволяют клеткам образовывать биопленку (которая увеличивает патогенную силу организма и предотвращает проникновение антибиотиков). См. Рисунок выше о роли чувствительности кворума с двумя компонентами в образовании бактериальной биопленки.
Повторяющаяся логика дизайна
Я описал здесь только несколько примеров регуляторных систем с двумя компонентами, среди которых можно было бы привести много других примеров. Благодаря этим различным системам мы наблюдаем повторяющуюся логику дизайна, несмотря на то, что эти системы не связаны эволюционными потомками. Это именно то, что мы могли бы ожидать, если бы тот же интеллектуальный дух был вовлечен в их происхождение, но на самом деле это довольно удивительно, если мы начнем с постулата унисона случайной и физической необходимости.


